
Predatory Functional Response of Harmonia sedecimnotata to Aphis craccivora
-
摘要:目的
明确纤丽瓢虫(Harmonia sedecimnotata)对豆蚜(Aphis craccivora)的捕食能力,为纤丽瓢虫田间防控豆蚜的应用提供理论依据。
方法以纤丽瓢虫和豆蚜为研究对象,设置不同虫态纤丽瓢虫与不同密度梯度的高龄豆蚜组合,采用室内捕食的方法,结合数据模型拟合,明确纤丽瓢虫各虫态对豆蚜的捕食功能反应、搜寻效应和自身密度干扰反应。
结果纤丽瓢虫1~4龄幼虫和雌雄成虫对豆蚜的捕食功能反应均符合Holling Ⅱ模型,捕食量随猎物密度增加而增加,且增加速率逐渐减小;1~4龄幼虫和雌雄成虫对豆蚜的瞬时攻击率分别为0.824、0.808、1.364、1.139、1.499、1.626,处理时间分别为0.060、0.014、0.006、0.003、0.004、0.005 d,日最大捕食量分别为16.667、71.428、166.667、333.333、250.000、200.000头;纤丽瓢虫1~4龄幼虫和成虫的捕食能力由强到弱分别为4龄幼虫、雌成虫、雄成虫、3龄幼虫、2龄幼虫和1龄幼虫;幼虫捕食量随龄期升高而增加,4龄幼虫的单头猎物处理时间最短(0.003 d)、捕食能力最强(379.667)、日最大捕食量最高(333.333头)。纤丽瓢虫对豆蚜的搜寻效应随猎物密度增加而降低,雌成虫的搜寻效应均大于雄成虫。纤丽瓢虫自身密度对豆蚜的捕食作用影响符合Hassell模型,雌雄成虫个体间存在种内干扰,干扰方程分别为E=0.465P-0.515和E=0.366P-0.505,平均捕食效率随猎物密度增加而降低,分摊竞争强度随猎物密度增加而升高,平均单头捕食量随捕食者密度增大而减少,当捕食者密度达5头/ 皿时,纤丽瓢虫雌雄成虫捕食效率最低、分别为0.198和0.172,分摊竞争强度最大、分别为0.548和0.558。
结论纤丽瓢虫对豆蚜具有较强的捕食能力,是豆蚜生物防治的重要天敌昆虫,在实际应用中应根据豆蚜发生密度,采用适宜的纤丽瓢虫虫态和数量进行投放。
Abstract:ObjectiveThe study was conducted to clarify the predation capacity of Harmonia sedecimnotata on Aphis craccivora, with a view to providing a theoretical basis for field control of A. craccivora by H. sedecimnotata.
MethodH. sedecimnotata and A.craccivora were taken as experimental subjects. H. sedecimnotata at different stages and different density gradients of late instar nymphs of A.craccivora were prepared to determine the predatory functional response, search effect and intraspecific interference response of H.sedecimnotata to A.craccivora by indoor predation method and numerical model.
ResultThe predatory functional response of 1-4th instar of larvae and adults of H. sedecimnotata conformed to Holling Ⅱ model. The predation amount increased with the increase of prey density. The instant attack rates (a) of 1-4st larvae, female and male adults were 0.824, 0.808, 1.364, 1.139, 1.499, 1.626, the handling time (Th) were 0.060, 0.014, 0.006, 0.003, 0.004, 0.005 d, and the maximum daily predation amounts were 16.667, 71.428, 166.667, 333.333, 250.000 and 200.000, respectively. The predation capacity of 1-4th instar larvae and adults of H. sedecimnotata ranked as (from strong to weak): 4th instar, female adult, male adult, 3rd instar, 2nd instar and 1st instar, respectively. The predation amount of larvae increased with the increase of age. For the 4th larvae, the handling time (Th) was the shortest, the predation capacity(a/Th) was the highest, and the maximum daily predation was the highest. which were 0.003 d, 379.667 and 333.333 aphids, respectively. The search effect of H. sedecimnotata on A.craccivora was decreased with the increase of prey density. The search effect of female was greater than that of male. The effect of the H. sedecimnotata's density on the predatory effect of A. craccivora was consistent with the Hassell model. There was intraspecific interference among adult H.sedecimnotata, and the interference equations of female and male adults were E=0.465P-0.515 and E=0.366P-0.505, respectively. The average predation rate decreased with the increase of prey density, the intensity of scramble competition increased with the increase of prey density, and the prey killed by per predator decreased with the increase of predator density. When the predator density reached 5 individuals per plate, the predation rates of female and male adults was the lowest, which were 0.198 and 0.172, and the intensity of scramble competition was the highest, which were 0.548 and 0.558, respectively.
ConclusionH. sedecimnotata, with a strong predatory ability on the A. craccivora, is an important natural enemy in the biological control of A. craccivora. In practical application, suitable insect state and quantity should be adopted according to the occurrence density of A. craccivora.
-
【研究意义】纤丽瓢虫(Harmonia sedecimnotata)为瓢虫科(Ccocinelliae)和谐瓢虫属(Harmonia)昆虫,广泛分布于南亚和东南亚地区,在我国主要分布于西南和华南地区[1],常见于广州田间、公园、湿地等环境,为本地优势种类,可捕食桃蚜(Myzus persicae)、棉蚜(Aphis gossypii)[2-3]。田间调查发现,纤丽瓢虫还可捕食紫薇长斑蚜和豆蚜(Aphis craccivora)。捕食功能反应是研究捕食者对猎物捕食潜能的重要工具,揭示捕食者与猎物之间的动态关系[4]。Holling[5]提出3种捕食功能反应模型,即Holling Ⅰ、Ⅱ、Ⅲ模型,分别通过直线回归、曲线回归和“S”型曲线描述捕食者对猎物不同密度的捕食率、攻击系数和处理时间等,以明确捕食者对猎物的捕食潜能[6]。其中,最能反映节肢动物捕食功能的模型为Holling Ⅱ和Ⅲ[7]。因此,通过测定捕食功能反应,明确纤丽瓢虫捕食豆蚜的潜能,对纤丽瓢虫防控豆蚜的实践应用具有重要意义。【前人研究进展】生物防治具有安全、环保、低能耗等优点,对降低化学农药使用、实现农业可持续发展具有重要意义[8]。捕食性瓢虫是生物防治的一类重要天敌昆虫,对控制蚜虫、介壳虫、粉虱、叶螨等农林害虫具有重要作用[1, 9]。全球已记录瓢虫6 896种、中国记录1 072种[10],表明瓢虫资源极为丰富。在我国,研究和应用较多的瓢虫种类有龟纹瓢虫(Propylaea japonica)、异色瓢虫(Harmonia axyridis)、七星瓢虫(Coccinella septempunctata)、孟氏隐唇瓢虫(Cryptolaemus montrouzieri)、六斑月瓢虫(Menochilus sexmaculata)等[11-15]。Boopathi等[2]通过对纤丽瓢虫生物学特性及其对棉蚜的捕食能力、田间应用评价研究,发现纤丽瓢虫具有良好的生物防治潜能。Semyanov等[3]研究了纤丽瓢虫取食桃蚜和麦蛾(Sitotroga cerealella)卵的寿命、繁殖力和捕食量等生物学特征,发现桃蚜更利于纤丽瓢虫生长繁殖。Reznik等[16]发现纤丽瓢虫的营养滞育诱导和终止受脂肪变化的影响,蚜虫丰富度与脂肪含量显著相关。豆蚜是重要的农业害虫,可为害蚕豆、豇豆、土豆、甜菜、辣椒等,导致植物叶片失绿、卷曲,蚜虫密度过高时甚至导致植株枯萎,同时豆蚜分泌的蜜露可诱发真菌病害煤污病,导致叶表面片发黑,降低植物光合作用,严重影响植物生长[17-18]。长期以来,规模化防治豆蚜的手段依赖化学农药,如阿维菌素、拟除虫菊酯类、氟啶虫酰胺、吡虫啉等,但上述药物残留在土壤、水体或飘散在空中,对环境和人体造成潜在危害,且可危害其他非靶标生物,对生物多样性造成负面影响[19-20]。长期使用化学农药可能增强蚜虫抗药性,进而减弱农药对蚜虫的防治效果[20]。因此,使用天敌昆虫防治豆蚜成为研究和应用的焦点。潘明真等[21]综述了近60年来我国豆蚜生物防治研究与应用成果,发现豆蚜的天敌昆虫非常丰富,包括隐蚜茧蜂(Aphidius funebris)、六斑月瓢虫(Menochilus sexmaculata)、大草蛉(Chrysopa pallens)、东亚小花蝽(Orius sauteri)、黑带食蚜蝇(Episyrphus balteatus)和食蚜瘿蚊(Aphidoletes aphidimyza)等,许多优势天敌昆虫正在被研究或投入应用,如七星瓢虫、烟蚜茧蜂(Aphidius gifuensis)、大草岭(Chrysopa pallens)等。研究发现,利用茶足柄瘤蚜茧蜂(Lysiphlebus testaceipes)和多异瓢虫(Hippodamia variegate)能较好控制田间豆蚜为害[22-23],印楝素和苦参碱对豆蚜也有较好防效[24-25]。【本研究切入点】为在生物防治应用方面建立理论支撑,多种常见优势天敌瓢虫在生物学、生态学、捕食功能反应、抗性等方面具有较为系统的研究,如龟纹瓢虫、异色瓢虫、七星瓢虫、六斑月瓢虫[11-13, 15]。纤丽瓢虫具有良好的生物防治应用潜能,捕食功能反应是应用评价的重要内容,而有关纤丽瓢虫对豆蚜的捕食功能研究尚未见报道。【拟解决的关键问题】本研究通过Holling Ⅱ模型和自身密度干扰反应模型构建纤丽瓢虫对豆蚜的捕食功能反应方程,明确纤丽瓢虫对豆蚜的捕食潜能、搜寻效应和自身密度竞争强度。
1. 材料与方法
1.1 试验材料
本试验于2024年8月在广州市林业和园林科学研究院植物保护所实验室完成。纤丽瓢虫采自广州流花湖公园紫薇(Lagerstroemia parviflora)植株,室内使用蚕豆苗繁殖的豆蚜饲养多代,饲养条件为温度26(±1)℃、相对湿度70(±5)%、光周期为L∶D=14∶10。
豆蚜准备:采用水培方式培育蚕豆苗,当蚕豆苗长至株高约5 cm时接种豆蚜,再剪下长满豆蚜的蚕豆苗置于新长成的蚕豆苗上,饲养条件同上,豆蚜繁殖1周后,用于饲喂瓢虫。
1.2 试验方法
1.2.1 纤丽瓢虫对豆蚜的捕食功能反应
收集12 h内纤丽瓢虫产的卵,于室温下自然孵化。待卵孵化后,使用带蚕豆苗的豆蚜饲喂,每天更换新鲜蚜虫,直至羽化为成虫。随机挑取羽化5 d内的成虫单头置于培养皿中饥饿处理24 h;随机挑取蜕皮12 h内的2~4龄幼虫饥饿处理12 h,1龄幼虫孵化8 h内即用于试验。将高龄(3~4龄)豆蚜若虫接入培养皿(直径9 cm、高1.5 cm)中,纤丽瓢虫1~4龄幼虫和成虫对应的豆蚜密度梯度分别为10、20、30、40、50头/皿,20、40、60、80、100头/皿,40、60、80、100、120头/皿,40、70、100、130、150、180头/皿,70、100、130、160、190、220头/皿。分别将上述纤丽瓢虫1~4龄幼虫和成虫单头饲养在对应豆蚜密度的培养皿中,每个密度梯度5次重复,试验环境条件同1.1。待纤丽瓢虫取食24 h后,统计蚜虫存活数量,计算捕食量。纤丽瓢虫对豆蚜的捕食功能反应采用Holling Ⅱ模型[2]进行拟合,计算公式如下:

式中,Na为捕食量,a为瞬时攻击率,N0为猎物密度,Th为处理时间(捕食单头猎物所需时间),T为猎物暴露于捕食者的时间(本试验中T=1 d),a/Th为捕食能力(比值越大,表示捕食能力越强),1/Th为日最大捕食量。将上述方程取倒数进行变形,即

采用最小二乘法即可求得a和Th。
1.2.2 纤丽瓢虫对豆蚜的搜寻效应
搜寻效应指天敌在捕食过程中对猎物的寻找行为,猎物密度越大,寻找时间越短,捕食效率越高。基于1.2.1的捕食功能反应方程,可得搜寻效应计算公式:

式中,a为攻击系数,Th为处理时间,N为猎物密度。
1.2.3 纤丽瓢虫自身密度干扰反应
随机挑取羽化5 d内的纤丽瓢虫成虫单头置于培养皿饥饿24 h。将高龄(3~4龄)豆蚜若虫接入培养皿,每皿300头,再接入饥饿处理后的纤丽瓢虫,雌雄成虫均设置5个密度梯度,分别为1、2、3、4、5头/皿,每个密度梯度5次重复,试验环境条件同1.1。待瓢虫取食24 h后,统计蚜虫存活数量,计算瓢虫捕食量。纤丽瓢虫自身密度干扰反应采用Hassell and Varley干扰模型[26]进行拟合,计算公式如下:

式中,E为捕食效率,利用公式E=Na/N0P求得;Q为搜寻常数,P为捕食者密度,m为干扰系数。
纤丽瓢虫自身密度干扰反应中,单个捕食者对猎物产生的竞争为分摊竞争[26],随着捕食者密度增加,分摊竞争强度发生变化。分摊竞争强度公式如下:

式中,E1为一头捕食者的捕食率,Ep为P头捕食者的捕食率。
1.3 数据处理与分析
原始数据采用Excel 2010整理,使用SPSS 21.0对数据进行回归分析和线性拟合。
2. 结果与分析
2.1 纤丽瓢虫对豆蚜的捕食功能反应
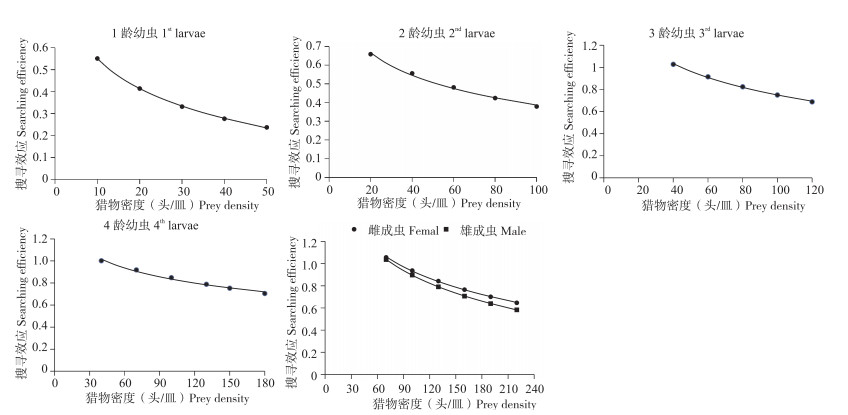
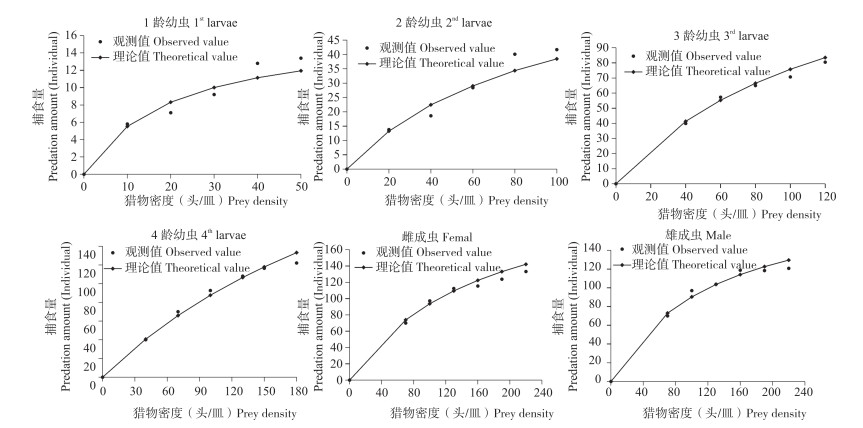
纤丽瓢虫的捕食量随猎物密度增加而增加,但增加速率逐渐减小,当猎物密度达到一定量后,捕食量变化极小,直至达到最大捕食量(图 1)。采用Holling Ⅱ模型得出纤丽瓢虫1~4龄幼虫和成虫对应的捕食功能反应方程及参数见 表 1,所有线性回归的R2均大于0.886,表明拟合度较高,捕食功能反应方程能较好反映纤丽瓢虫对豆蚜的捕食关系。通过模型分析,计算出纤丽瓢虫对豆蚜的瞬时攻击率、处理时间、捕食能力和日最大捕食量。纤丽瓢虫1~4龄幼虫、雌雄成虫对猎物的处理时间分别是0.060、0.014、0.006、0.003、0.004、0.005 d,捕食能力分别为13.733、57.714、227.333、379.667、374.750、325.200,日最大捕食量分别为16.667、71.428、166.667、333.333、250.000、200.000头。纤丽瓢虫1~4龄幼虫日最大捕食量随龄期增加而增加,雌成虫的日最大捕食量高于雄成虫。成虫的瞬时攻击率均高于幼虫,其中以雄成虫最强。纤丽瓢虫幼虫龄期越高,对单头猎物的处理时间越短,4龄幼虫的处理时间仅为0.003 d,低于成虫处理时间。纤丽瓢虫捕食能力由强到弱分别是4龄幼虫、雌成虫、雄成虫、3龄幼虫、2龄幼虫和1龄幼虫。4龄幼虫的日最大捕食量最高、捕食能力最强;1龄幼虫的日最大捕食量最小、捕食能力最弱。捕食功能反应方程显示,纤丽瓢虫3~4龄幼虫和成虫对豆蚜均有较好的防治潜能。
![]() 图 1 不同虫态纤丽瓢虫对豆蚜的捕食功能反应曲线Figure 1. Predatory functional response curves of Harmonia sedecimnotata to Aphis craccivora表 1 不同虫态纤丽瓢虫对豆蚜的捕食功能反应方程及参数Table 1. Predatory functional response equation and parameters of Harmonia sedecimnotata to Aphis craccivora
图 1 不同虫态纤丽瓢虫对豆蚜的捕食功能反应曲线Figure 1. Predatory functional response curves of Harmonia sedecimnotata to Aphis craccivora表 1 不同虫态纤丽瓢虫对豆蚜的捕食功能反应方程及参数Table 1. Predatory functional response equation and parameters of Harmonia sedecimnotata to Aphis craccivora
2.2 纤丽瓢虫对豆蚜的搜寻效应
搜寻效应揭示了捕食者与猎物之间的攻击行为,其与猎物密度和捕食者本身的攻击能力有关。纤丽瓢虫对高龄豆蚜若虫的搜寻效应随着猎物密度增加而降低,表明在高猎物密度条件下,纤丽瓢虫捕食豆蚜所用时间减少、效率增加(图 2)。纤丽瓢虫4龄幼虫的搜寻效应下降趋势较缓慢,1龄幼虫和成虫的搜寻效应下降趋势较快,表明猎物密度对1龄幼虫和成虫的攻击行为影响更大。纤丽瓢虫雌雄成虫对应6个蚜虫密度梯度的搜寻效应分别为1.056、0.937、0.842、0.765、0.700、0.646, 1.036、0.897、0.791、0.707、0.639、0.583,在相同密度条件下,纤丽瓢虫雌成虫的搜寻效应均大于雄成虫,意味着雌成虫可能需要消耗更长的时间捕食同等数量的猎物。
![]() 图 2 不同虫态纤丽瓢虫对豆蚜的搜寻效应Figure 2. Search effects of Harmonia sedecimnotata to Aphis craccivora
图 2 不同虫态纤丽瓢虫对豆蚜的搜寻效应Figure 2. Search effects of Harmonia sedecimnotata to Aphis craccivora2.3 纤丽瓢虫成虫自身密度干扰反应
经干扰反应模型拟合,结果(表 2)显示雄雌成虫干扰方程的R2均大于0.9,表示方程拟合度较高,能够体现纤丽瓢虫对豆蚜的自身密度干扰反应。纤丽瓢虫成虫平均单头捕食量随着捕食者密度增大而减小,当雌雄成虫密度为5头/皿时,平均单头捕食量最低、分别为59.267头和51.333头;平均捕食效率随着猎物密度增加而降低,分摊竞争强度则反之。在同等捕食者密度条件下,雄虫的平均捕食效率低于雌虫,雄虫的分摊竞争强度高于雌虫。当捕食者密度达5头/皿时,捕食效率最低,分摊竞争强度最大。
表 2 纤丽瓢虫成虫对豆蚜的自身密度干扰反应参数Table 2. Intraspecific interference parameters of different densities of Harmonia sedecimnotata to Aphis craccivora
3. 讨论
本研究通过室内捕食试验分析纤丽瓢虫对豆蚜的捕食量、捕食功能反应、搜寻效应和自身密度干扰反应,系统评价纤丽瓢虫对豆蚜的生物防治潜能。结果显示,纤丽瓢虫1~4龄幼虫和成虫均能较好拟合Holling Ⅱ模型,这与七星瓢虫[27]、六斑月瓢虫[28]、异色瓢虫[29]、十斑大瓢虫(Megalocaria dilatata)[30]、龟纹瓢虫[31]、东亚小花蝽(Orius sauteri)[32]、食蚜蝇(Allograpta exotica)[33]、大草岭[34]、食蚜瘿蚊[35]对豆蚜以及红彩真猎蝽对斜纹夜蛾[36]的捕食功能反应模型一致。表明Holling Ⅱ模型能够反映多种天敌对不同猎物的捕食功能特征,在使用上述天敌昆虫开展生物防治时应采用淹没式的释放方法[37]。
温度对瓢虫的捕食能力有显著影响[38]。例如,异色瓢虫在15℃时对豌豆蚜(Acyrthosiphon pisum)的捕食能力最强[39];七星瓢虫和长足瓢虫(Hippodamia convergens)的捕食量随温度升高而增加,在23℃时对桃蚜的捕食量最大[40];菱斑巧瓢虫(Oenopia conglobata)在25℃时对木虱(Agonoscena pistaciae)的瞬时攻击率和日最大捕食量均高于22℃,且25℃的处理时间少于22℃[41]。本研究对纤丽瓢虫的捕食功能试验在26(±1)℃条件进行,可能不是最佳的捕食温度。未来将开展不同温度条件下纤丽瓢虫对豆蚜的捕食功能反应研究,以明确纤丽瓢虫的最佳捕食温度和不同温度条件下捕食参数发展趋势,为不同季节和气温条件下田间定量释放纤丽瓢虫提供理论参考。研究表明,纤丽瓢虫对棉蚜的捕食效率随着发育阶段升高而增加,4龄是捕食量最大的幼虫阶段,日最大捕食量为200.4头;成虫的捕食效率最高,其中雌成虫的捕食量大于雄成虫[15]。六斑月瓢虫和龟纹瓢虫对柑橘木虱的最大捕食能力和最高捕食量均出现在4龄幼虫期,其次是成虫期[42-43];六斑月瓢虫对棉蚜的日最大捕食能力最强的是4龄幼虫期,日最大捕食量为96.1头,且雄成虫与4龄幼虫的捕食量差异不显著[3]。十斑大瓢虫对豆蚜的瞬间攻击率和日最大捕食量均出现在4龄幼虫期,分别为4.7146和988.1128头[30]。七星瓢虫4龄幼虫对豌豆修尾蚜日捕食量最大、为625.0头,该龄期幼虫的单头猎物处理时间最短、为0.0016 d[12]。本研究发现纤丽瓢虫对豆蚜的最高捕食能力和最大捕食量发生在4龄幼虫期、分别为379.667和333.333头,其次为成虫期,推测4龄幼虫是瓢虫的取食高峰期,期间发育速度快,需储藏大量营养物质,为化蛹作准备。因此,在使用天敌瓢虫进行田间控害时,应根据蚜虫种群密度选择适合的虫态:在蚜虫发生盛期、种群密度较高时,宜采用高龄幼虫和成虫;在蚜虫发生初期、种群密度较低时,可采用低龄幼虫或者卵。
许多天敌瓢虫对猎物的搜寻效应随着猎物密度增加而降低,如七星瓢虫、龟纹瓢虫和六斑月瓢虫等[12, 42-43]。本研究发现纤丽瓢虫对豆蚜的搜寻效应随着蚜虫密度增加而出现缓慢下降的趋势,且1龄幼虫和成虫的下降趋势更明显,表明当蚜虫密度升高时,纤丽瓢虫捕食蚜虫所用的时间减少、捕食率增加,1龄幼虫和成虫的捕食效应对豆蚜密度变化反应相对滞缓。在相同猎物密度条件下,纤丽瓢虫雌成虫的搜寻效应均大于雄成虫,这与六斑月瓢虫对豆蚜捕食的结果[44]一致;瓢虫幼虫随着龄期增加搜寻效应也增大,如七星瓢虫对豌豆修尾蚜(Megoura crassicauda)的捕食作用[12],表明雌成虫和高龄幼虫需要消耗更多时间寻找猎物。捕食者自身密度对其捕食作用会产生强烈干扰。本研究结果表明,纤丽瓢虫对豆蚜的捕食存在较强的种内干扰效应,当蚜虫密度一定时,随着纤丽瓢虫密度增加,其捕食效率逐渐减小、分摊竞争强度加大,平均单头纤丽瓢虫的捕食量逐渐下降。在六斑月瓢虫捕食柑橘木虱[43]、七星瓢虫捕食豌豆修蚜[12]中也存在很强的种内干扰作用。在瓢虫密度相同的情况下,雌成虫的竞争强度均高于雄成虫,且雌成虫的食量大于雄成虫,可能是雌成虫需要产卵繁殖后代的原因。因此,田间释放纤丽瓢虫防控蚜虫时,应根据蚜虫种群数量控制天敌释放量,避免同时高密度释放,应采取多次分散释放的策略。
本研究在室内控制条件下进行,设置了较为适宜的温度、湿度和光照,且无其他干扰因素。结果表明,纤丽瓢虫对豆蚜具有较高的捕食量和捕食效率,室内观察发现纤丽瓢虫单一取食豆蚜能完成世代发育并繁殖后代。但天敌昆虫的应用效果受复杂的自然环境影响,包括天气情况、猎物种类、害虫寄主植物、天敌本身密度干扰等[45]。因此,仅将室内研究结果作为评价天敌实际应用的指标具有一定局限性,未来将结合田间开放试验,多元化全面评价天敌应用潜能。
4. 结论
纤丽瓢虫自身密度对豆蚜的捕食作用影响符合Holling Ⅱ模型,各龄期幼虫和成虫对豆蚜具有良好的捕食能力,捕食量随着猎物密度增加而增加,当猎物无限增加时,捕食量增加速率趋于平缓。纤丽瓢虫1~4龄幼虫和成虫的捕食能力由强至弱分别为4龄幼虫(379.667)、雌成虫(374.750)、雄成虫(325.200)、3龄幼虫(227.333)、2龄幼虫(57.714)和1龄幼虫(13.733)。幼虫捕食量随着龄期增加而增加,4龄幼虫的单头猎物处理时间最短(0.003 d)、捕食能力最强(379.667)、日最大捕食量最高(333.333头)。纤丽瓢虫对豆蚜的搜寻效应随着猎物密度增加而降低,表明猎物密度增加减少了捕食者的捕食时间。在相同密度条件下,雌成虫的搜寻效应大于雄成虫。纤丽瓢虫成虫个体间存在种内干扰,种内竞争强度随着天敌密度增大而增强,当捕食者密度达5头/皿时,捕食效率最低,分摊竞争强度最大。实际应用中可根据蚜虫密度调整瓢虫释放虫态和释放量,采用多次分散释放的方式。
【责任编辑 陈丽娥】 -
![]()
图 1 不同虫态纤丽瓢虫对豆蚜的捕食功能反应曲线
Figure 1. Predatory functional response curves of Harmonia sedecimnotata to Aphis craccivora
![]()
图 2 不同虫态纤丽瓢虫对豆蚜的搜寻效应
Figure 2. Search effects of Harmonia sedecimnotata to Aphis craccivora
表 1 不同虫态纤丽瓢虫对豆蚜的捕食功能反应方程及参数
Table 1 Predatory functional response equation and parameters of Harmonia sedecimnotata to Aphis craccivora
 下载: 导出CSV
下载: 导出CSV
表 2 纤丽瓢虫成虫对豆蚜的自身密度干扰反应参数
Table 2 Intraspecific interference parameters of different densities of Harmonia sedecimnotata to Aphis craccivora
下载: 导出CSV
-
[1] 任顺祥, 王兴民, 庞虹, 彭正强, 曾涛. 中国瓢虫原色图鉴[M]. 北京: 科学出版社, 2009: 2-19. REN S X, WANG X M, PANG H, PENG Z Q, ZENG T. Colored pictorial handbook of ladybird beetles in China[M]. Beijing: Science Press, 2009: 2-19.
[2] BOOPATHI T, SINGH S B, DUTTA S K, DAYAl V, SING A R, CHOUDHURY S, RAMAKRISHNA Y, ARAVINTHARAJ R, SHAKUNTALA I, LALHRUAIPUⅡ K. Harmonia sedecimnotata (F.): Predatory potential, biology, life table, molecular characterization, and feld evaluation against Aphis gossypii Glover[J]. Scientific Reports, 2020, 10(1): 3079. DOI: 10.1038/s41598-020-59809-3.
[3] SEMYANOV V P, VAGHINA N P. Effect of the trophic diapause on the fecundity and longevity of a tropical coccinellid, Harmonia sedecimnotata (Fabr.) (Coleoptera, Coccinellidae)[J]. Entomological Review, 2003, 83(8): 915-916.
[4] FENLON J S, FADDY M J. Modelling predation in functional response[J]. Ecological Modelling, 2006, 198(1): 154-162. DOI: 10.1016/j.ecolmodel.2006.04.002.
[5] HOLLING C S. The functional response of invertebrate predators to prey density[J]. The Memoirs of the Entomological Society of Canada, 1966, 98: 85-86. DOI: 10.4039/entm9848fv.
[6] VENKANNA Y, SUROSHE S S, CHANDER S, KUMARI S. Feeding potential and foraging behaviour of Cheilomenes sexmaculata (F.) on the cotton aphid, Aphis gossypii Glover[J]. International Journal of Tropical Insect Science, 2021, 41(4): 2431-2442. DOI: 10.1007/S42690-020-00420-4.
[7] HUFFAKER C B, RABB R L. Ecological entomology[M]. New York: Wiley Interscience, 1984: 497-530.
[8] 张礼生, 陈红印. 生物防治作用物研发与应用的进展[J]. 中国生物防治学报, 2014, 30(5): 581-586. DOI: 10.16409/j.cnki.2095-039x.2014.05.009. ZHANG L S, CHEN H Y. Advances in research and application of biological control agents in China[J]. Chinese Journal of Biological Control, 2014, 30(5): 581-586. DOI: 10.16409/j.cnki.2095-039x.2014.05.009.
[9] 王兴民, 陈晓胜, 邱宝利, 任顺祥. 捕食性瓢虫采集与调查取样技术[J]. 应用昆虫学报, 2014, 51(5): 1362-1366. DOI: 10.7679/j.issn.2095-1353.2014.162. WANG X M, CHEN X S, QIU B L, REN S X. Methods and technologies for collecting, surveying and sampling predatory ladybirds (Coleoptera: Coccinellidae)[J]. Chinese Journal of Applied Entomology, 2014, 51(5): 1362-1366. DOI: 10.7679/j.issn.2095-1353.2014.162.
[10] 王兴民, 陈晓胜. 中国瓢虫图鉴[M]. 福州: 海峡出版发行集团, 2022: 1-30. WANG X M, CHEN X S. Illustrated handbook of ladybird beetles in China[M]. Fuzhou: The Straits Publishing & Distributing Group, 2022: 1-30.
[11] 刘志伟, 钟国辉, 王永跃, 王兴民, 毕可可. 龟纹瓢虫生殖系统及卵子发生观察[J]. 广东农业科学, 2021, 48(6): 101-106. DOI: 10.16768/j.issn.1004-874X.2021.06.014. LIU Z W, ZHONG G H, WANG Y Y, WANG X M, BI K K. Reproductive system and oogenesis of Propylea japonica[J]. Guangdong Agricultural Sciences, 2021, 48(6): 101-106. DOI: 10.16768/j.issn.1004-874X.2021.06.014.
[12] KOCH R L, COSTAMAGNA A C. Reaping benefits from an invasive species: Role of Harmonia axyridis in natural biological control of Aphis glycines in North America[J]. BioControl, 2017, 62(3): 331-340. DOI: 10.1007/s10526-016-9749-9.
[13] 吴沐秀, 张晓媛, 昝庆安, 张宏瑞. 七星瓢虫对豌豆修尾蚜的捕食功能反应[J]. 南方农业学报, 2022, 53(11): 3128-3136. DOI: 10.3969/j.issn.2095-1191.2022.11.013. WU M X, ZHANG X Y, ZAN Q A, ZHAGN H R. Predatory functional response of Coccinella septempunctata to Megoura crassicauda[J]. Journal of Southern Agriculture, 2022, 53(11): 3128-3136. DOI: 10.3969/j.issn.2095-1191.2022.11.013.
[14] LI H S, HUANG Y H, CHEN M L, REN Z, QIU B Y, PATRICK D C, GERALD H, HONG P. Genomic insight into diet adaptation in the biological control agent Cryptolaemus montrouzieri[J]. BMC Genomics, 2021, 22: 135. DOI: 10.1186/S12864-021-07442-3.
[15] 唐良德, 李飞, 赵海燕, 吴建辉, 任顺祥. 六斑月瓢虫取食不同猎物的生长发育及繁殖特性[J]. 生态学报, 2017, 37(17): 5765-5770. DOI: 10.5846/stxb201606111117. TANG L D, LI F, ZHAO H Y, WU J H, REN S X. Influence of three prey species on the development and fecundity of Menochilus sexmaculata Fabricius[J]. Acta Ecologica Sinica, 2017, 37(17): 5765-5770. DOI: 10.5846/stxb201606111117.
[16] REZNIK S Y, VAGHINA N P. Dynamics of fat content during induction and termination of "trophic diapause" in Harmonia sedecimnotata Fabr. females (Coleoptera, Coccinellidae)[J]. Entomological Review, 2006, 86(2): 125-132. DOI: 10.1134/S0013873806020011.
[17] MANDOUR N S, NESREN A S B, LIU T X. Functional response of the ladybird, Cydonia vicina Nilotica to cowpea aphid, Aphis craccivora in the laboratory[J]. Insect Science, 2006, 13(1): 49-54. DOI: 10.1111/j.1744-7917.2006.00067.x.
[18] 唐良德, 王晓双, 赵海燕, 付步礼, 邱海燕, 刘奎. 大草蛉幼虫捕食豆大蓟马和豆蚜的功能反应及生长发育[J]. 中国生物防治学报, 2017, 33(1)49-55. DOI: 10.16409/j.cnki.2095-039x.2017.01.008. TANG L D, WANG X S, ZHAO H Y, FU B L, QIU H Y, LIU K. The predation function response and development of Chrysopa pallens larva on Megalurothrips usitatus and Aphis craccivora[J]. Chinese Journal of Biological Control, 2017, 33(1): 49-55. DOI: 10.16409/j.cnki.2095-039x.2017.01.008.
[19] ZALLER J G, MAREN K P, ULRICH S, EDITH G, MARIA P, IMRAN N, HERBERT F, PETER H H, LUKAS L. Pesticides in ambient air, influenced by surrounding land use and weather, pose a potential threat to biodiversity and humans[J]. The Science of the Total Environment, 2022, 838(2): 156012-156012. DOI: 10.1016/J.SCITOTENV.2022.156012.
[20] 汤秋玲, 马康生, 高希武. 蔬菜蚜虫抗药性现状及抗性治理策略[J]. 植物保护, 2016, 42(6): 11-20. DOI: 10.3969/j.issn.0529-1542.2016.06.002. TANG Q L, MA K S, GAO X W. Current status and management strategies of insecticide resistance in aphids on the vegetable crops[J]. Plant Protection, 2016, 42(6): 11-20. DOI: 10.3969/j.issn.0529-1542.2016.06.002.
[21] 潘明真, 张毅, 曹贺贺, 王杏杏, 刘同先. 我国主要农作物蚜虫生物防治的研究进展、应用与展望[J]. 植物保护学报, 2022, 49(1): 146-172. DOI: 10.13802/j.cnki.zwbhxb.2022.2022820. PAN M Z, ZHANG Y, CAO H H, WANG X X, LIU T X. Research progresses, application, and prospects in aphid biological control on main crops in China[J]. Journal of Plant Protection, 2022, 49(1): 146-172. DOI: 10.13802/j.cnki.zwbhxb.2022.2022820.
[22] 郭佩佩, 帕提玛·乌木尔汗, 任豪辉, 图尔荪阿依·艾散, 马德英. 释放不同益害比多异瓢虫对设施豇豆豆蚜的防效及定殖影响[J]. 中国生物防治学报, 2021, 38(2): 312-320. DOI: 10.16409/j.cnki.2095-039x.2021.05.002. GUO P P, WUMUERHAN P, REN H H, AISAN T, MA D Y. Influence of releasing Hippodamia variegate at different natural enemy/pest ratios on the control and colonization of Aphids craccivora on cowpea in protected cultivation[J]. Chinese Journal of Biological Control, 2021, 38(2): 312-320. DOI: 10.16409/j.cnki.2095-039x.2021.05.002.
[23] ZUMOFFEN L, TAVELLA J, SIGNORINI M, SALVO A. Laboratory and field studies to evaluate the potential of an open rearing system of Lysiphlebus testaceipes for the control of Aphis craccivora in Argentina[J]. BioControl, 2016, 61(1): 23-33. DOI: 10.1007/s10526-015-9698-8.
[24] 戴爱梅, 丁志梅, 张海军, 陈先荣, 戴秋萍, 陆永跃. 印楝素与脂肪酸甲酯联合使用对豆蚜的防治效果[J]. 环境昆虫学报, 2022, 44(5): 1301-1307. DOI: 10.3969/j.issn.1674-0858.2022.05.25. DAI A M, DING Z M, ZHANG H J, CHEN X R, DAI Q P, LU Y Y. Evaluation of the control effect of azadirachtin and fatty acid methyl ester spray adjuvant on Aphis craccivora (Coch)[J]. Journal of Environmental Entomology, 2022, 44(5): 1301-1307. DOI: 10.3969/j.issn.1674-0858.2022.05.25.
[25] 吴聪, 陈镓豪, 朱思怡, 汪暖. 大棚枇杷上木虱发生规律及5种杀虫剂的防治效果[J]. 中国南方果树, 2024, 53(5): 91-94, 100. DOI: 10.13938/j.issn.1007-1431.20230353. WU C, CHEN J H, ZHU S Y, WANG N. Occurrence of psyllid on loquat in greenhouses and control effect of 5 insecticides[J]. South China Fruits, 2024, 53(5): 91-94, 100. DOI: 10.13938/j.issn.1007-1431.20230353.
[26] HASSELL M P, VARLEY G C. New inductive population model for insect parasites and its bearing on biological control[J]. Nature, 1969, 223(5211): 1133-1137. DOI: 10.1038/2231133a0.
[27] CHHANGANI G, MAHLA M K, SWAMINATHAN R, SWAMI H, DANGI N L, AGARWALA B K. Comparative functional and numeric response of two coccinellids (Coccinella septempunctata and Cheilomenes sexmaculata) preying cowpea aphid (Aphis craccivora)[J]. Legume Research-An International Journal, 2022, 45(4): 521-526. DOI: 10.18805/LR-4697.
[28] PATEL P A. Feeding potentiality of Menochilus sexmaculatus Fab. on different aphid species[J]. International Journal on Recent and Innovation Trends in Computing and Communication, 2015, 3(7): 4426-4430. DOI: 10.17762/ijritcc.v3i7.4666.
[29] LIU Y H, LI R, LI S G. Consumption patterns of the multicolored Asian ladybird Harmonia axyridis on the broad bean aphid Aphis craccivora[J]. Journal of Asia-Pacific Entomology, 2021, 25(1): 101852. DOI: 10.1016/J.ASPEN.2021.101852.
[30] 魏莲, 刘虹伶, 伍兴隆, 陈河竹, 彭应力, 肖科军, 蔡鹏, 房超, 李跃建, 蒲德强. 十斑大瓢虫对豆蚜的捕食特性研究[J]. 中国农学通报, 2024, 40(5): 105-109. DOI: 10.11924/j.issn.1000-6850.casb2023-0167. WEI L, LIU H L, WU X L, CHEN H Z, PENG Y L, XIAO K J, CAI P, FANG C, LI Y J, PU D Q. Study on the predation characteristics of Megalocaria dilatata on Aphid[J]. Chinese Agricultural Science Bulletin, 2024, 40(5): 105-109. DOI: 10.11924/j.issn.1000-6850.casb2023-0167.
[31] PERVEZ A. Functional responses of coccinellid predators: An illustration of a logistic approach[J]. Journal of Insect Science, 2005, 5(5): 1-6. DOI: 10.1673/031.005.0501.
[32] GE Y, CAMARA I, WANG Y, LIU P P, ZHANG L, XING Y J, LI A M, SHI W P. Predation of Aphis craccivora (Hemiptera: Aphididae) by Orius sauteri (Hemiptera: Anthocoridae) under different temperatures[J]. Journal of Economic Entomology, 2018, 111(6): 2599-2604. DOI: 10.1093/jee/toy255.
[33] ARCAYA E, BAÑÓN C P, MENGUAL X, VALLEJO J J Z, ROJO S. Life table and predation rates of the syrphid fly Allograpta exotica, a control agent of the cowpea aphid Aphis craccivora[J]. Biological Control, 2017, 115: 74-84. DOI: 10.1016/j.biocontrol.2017.09.009.
[34] ABDEL-HAMEID N F. Functional response estimations of Chrysoperla carnea to different densities of Aphis craccivora and Gynaikothrips fi corum nymphs[J]. Brazilian Journal of Biology, 2022, 84: e268066. DOI: 10.1590/1519-6984.268066.
[35] MADAHI K, SAHRAGARD A, HOSSEINI R. Predation rate and numerical response of Aphidoletes aphidimyza feeding on di f ferent densit ies of Aphi s craccivora[J]. Biocontrol Science and Technology, 2015, 25(1): 72-83. DOI: 10.1080/09583157.2014.948383.
[36] 邓海滨, 王珍, 陈永明, 吴文斌, 彭文松. 红彩真猎蝽对斜纹夜蛾和烟青虫的捕食功能反应[J]. 广东农业科学, 2012, 39(13): 107-109. DOI: 10.3969/j.issn.1004-874X.2012.13.036. DENG H B, WANG Z, CHEN Y M, WU W B, PENG W S. Predation of Harpactor fuscipes on Helicoverpa assulta and Spodoptera litura[J]. Guangdong Agricultural Sciences, 2012, 39(13): 107-109. DOI: 10.3969/j.issn.1004-874X.2012.13.036.
[37] LENTEREN J C V, HEMERIK L, LINS J C, BUENO V H P. Functional responses of three neotropical mirid predators to eggs of Tuta absoluta on tomato[J]. Insects, 2016, 7(3): 34. DOI: 10.3390/insects7030034.
[38] KHAN J, KHAN A, AHMED N, ALHAG S K, ALMADIY A A, SAYED S, ALAM P, ULLAH F. Age and stage-specific life table parameters of Harmonia dimidiata (Coleoptera: Coccinellidae) fed on Rhopalosiphum padi (Hemiptera: Aphididae) at different temperatures[J]. Egyptian Journal of Biological Pest Control, 2022, 32(1): 1-9. DOI: 10.1186/S41938-022-00610-X.
[39] YASIR I, ALI G, FAN Y D, ZHOU X M, AFIFA N, MAHMOOD S F. Age-stage, two-sex life table and predation parameters of Harmonia axyridis Pallas (Coleoptera: Coccinellidae), reared on Acyrthosiphon pisum (Harris) (Hemiptera: Aphididae), at four different temperatures[J]. Crop Protection, 2022, 159: 106029. DOI: 10.1016/J.CROPRO.2022.106029.
[40] KATSAROU I, MARGARITOPOULOS J T, TSITSIPIS J A, PERDIKIS D C, ZARPAS K D. Effect of temperature on development, growth and feeding of Coccinella septempunctata and Hippodamia convergens reared on the tobacco aphid, Myzus persicae nicotianae[J]. BioControl, 2005, 50(4): 565-588. DOI: 10.1007/s10526-004-2838-1.
[41] SANATI S, GOLDASTEH S, SHIRVANI A, RASHKI M. Functional response of Oenopia conglobata contaminata (Coleoptera: Coccinellidae) to Agonoscena pistaciae (Hemiptera: Psyllidae) at two different temperatures[J]. International Journal of Tropical Insect Science, 2020, 40(3): 621-628. DOI: 10.1007/s42690-020-00108-9.
[42] 毛加梅, 杨建东, 郭俊, 岳建强, 闫素云, 赵俊, 王自然. 捕食螨释放量对其防控柑桔全爪螨效果的影响[J]. 中国南方果树, 2024, 53(6): 37-42. DOI: 10.13938/j.issn.1007-1431.20230552. MAO J M, YANG J D, GUO J, YUE J Q, YAN S Y, ZHAO J, WANG Z R. Effect of predator mite release on the control effect of Panonychus mite in citrus[J]. South China Fruits, 2024, 53(6): 37-42. DOI: 10.13938/j.issn.1007-1431.20230552.
[43] 何万财, 叶青青, 王飞凤, 胡玉伟, 刘金华, 毛熊兴, 张瑞峰, 王兴民, 邱宝利, 刘玉涛, 桑文. 六斑月瓢虫对柑橘木虱的捕食作用[J]. 中国生物防治学报, 2023, 39(3): 514-522. DOI: 10.16409/j.cnki.2095-039x.2023.05.001. HE W C, YE Q Q, WANG F F, HU Y W, LIU J H, MAO X X, ZHANG R F, WANG X M, QIU B L, LIU Y T, SANG W. Predatory responses of Cheilomenes sexmaculata to Diaphorina citri[J]. Chinese Journal of Biological Control, 2023, 39(3): 514-522. DOI: 10.16409/j.cnki.2095-039x.2023.05.001.
[44] 唐良德, 李飞, 吴建辉, 任顺祥. 六斑月瓢虫捕食不同猎物的功能反应研究[J]. 中国生物防治学报, 2015, 31(2): 202-207. DOI: 10.16409/j.cnki.2095-039x.2015.02.007. TANG L D, LI F, WU J H, REN S X. Predation function response of Menochilus sexmaculata adults to Bemisia tabaci, Aphis craccivora and Rhopalosiphum maidis[J]. Chinese Journal of Biological Control, 2015, 31(2): 202-207. DOI: 10.16409/j.cnki.2095-039x.2015.02.007.
[45] LI D X, TIAN J, SHEN Z R. Functional response of the predator Scolothrips takahashii to hawthorn spider mite, Tetranychus viennensis: Effect of age and temperature[J]. BioControl, 2007, 52(1): 41-61. DOI: 10.1007/s10526-006-9015-7.
计量
- 文章访问数: 9
- HTML全文浏览量: 5
- PDF下载量: 3


